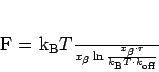The first step to understand interactions in cells and between cells is to understand interactions between the involved biomolecules. Besides DNA/RNA and antigen-antibody interactions, there are many proteins (receptors) and corresponding ligands that play a vital role inside and in-between cells.
0.5
![\includegraphics[width=.5\textwidth]{Bilder/streptavidin-bondforce}](img31.png)
|
Because of their exceptionally high binding affinity, two of
the most prominent ligand-receptor pairs are
streptavidin-biotin and avidin-biotin. Both proteins have a
tetrameric
structure, so they can bind up to four ligands.
Although many properties are valid for other ligand-receptor pairs,
only these two ligand-receptor pairs were used in this thesis and,
therefore, this section will focus on them.
During the last two decades, the rupture force between ligand and
receptor was investigated with several different methods, such as Atomic
Force Microscopy (AFM)
[44,97,86,122,28,134], Surface
Force Apparatus (SFA) [63,137], Dynamic Force Spectroscopy
(DFS) [39,91], Flow Chamber measurements
[102] and relaxation experiments with magnetic nanoparticles
[77]. Complementary to the measurements,
computer-
simulations were done by several
groups
[59,70,40,65]. Figure
1.4 shows a single streptavidin-biotin
pair and
exemplarily the applied
force in bond-force measurements.
Numerous measurements of the rupture force between ligand-receptor
pairs were made with the AFM. In AFM experiments, the tip is coated by
one part of the ligand-receptor pair and the other is fixed to a surface
(e.g. an agarose bead [44]). After the tip contacts the
surface and the ligand-receptor pair is bound, the tip is retracted
using the force spectroscopy mode. Under appropriate experimental
conditions [123,122], hundreds of single bond breaking
events can be measured within a short time. But AFM measurements are
somewhat limited, as shown in 1999 by MERKEL et al.
[91]. They showed that the bond-force depends strongly on
the rate of force increase ![]() 1.4. Although this dependency was found with AFM
measurements as well [134], only the DFS has the
possibility to measure with loading-rates from 0.05pN/sec to
60nN/sec. Dynamic Force Spectroscopy, developed by EVANS et al.
[39] in 1991, utilises biomembrane probes and is a potent
method for the analysis of rupture events. DFS consists of two
micropipettes that push two vesicles against each other. A vesicles
contains low amounts of either receptors or ligands. The separation of
the vesicles is analysed under a microscope, and video is recorded to
calculate the rupture force.
1.4. Although this dependency was found with AFM
measurements as well [134], only the DFS has the
possibility to measure with loading-rates from 0.05pN/sec to
60nN/sec. Dynamic Force Spectroscopy, developed by EVANS et al.
[39] in 1991, utilises biomembrane probes and is a potent
method for the analysis of rupture events. DFS consists of two
micropipettes that push two vesicles against each other. A vesicles
contains low amounts of either receptors or ligands. The separation of
the vesicles is analysed under a microscope, and video is recorded to
calculate the rupture force.
In contrast to the AFM measurements, where the measured bond-forces where around 100 to 300pN, it was possible to measure bond-forces of only 5pN with the DFS [91]. Experiments with other techniques, also got results in between those values, which support the loading-rate dependency. In addition, this thesis presents experiments that confirm the dependency in the range of extremely low loading-rates (see chapter 4).
In a different approach multi-wavelength x-ray diffraction methods were used to obtain the specific structure of ligand-receptor pairs with å ngstrøm precision [132,64,88,46,24]. Results of such x-ray diffraction methods can be seen in figures 1.2, 1.3 and 1.4.
In 1987 WEBER et al. [132] were the first group who
fabricated streptavidin crystals and characterised the
streptavidin-biotin complex with this method. Two years later
HENDRICKSON et al. [64] made a comprehensive assay
using a multi-wavelength anomalous diffraction method at an x-ray energy
of 11921eV. They found that the biotin is buried deeply inside the
![]() -barrel of the streptavidin protomer. Only the carboxylate
oxygens and the ureido-ring nitrogen protrude to the outside.
Multitudinous hydrogen-bonds and van der Waals interactions are involved
in the biotin binding. There are three hydrogen bonds to the carbonyl
group buried within the barrel and also hydrogen bonds to the ureido
nitrogens and carboxyl oxygens. Four tryptophan amino acids are in
contact with each biotin molecule. Most of the interactions result from
the residues of a given subunit, although one tryptophan is supplied by
a subunit related to the R diad axis, which is vital for the tetramer
integrity. This is an explanation for the reduced affinity with less
than four ligands that was also found elsewhere
[76,113]. WEBER et al. [131] confirmed
the results for streptavidin and apostreptavidin in 1992 and
LIVNAH et al. [88] got similar results for the
avidin-biotin complex in 1993. In 1998, CHU et al. [24]
presented a very convincing experiment, where they removed the
polypeptide loop that undergoes an open to closed conformational change
when biotin is bound. They showed that the deletion caused a large
decrease of the affinity for the full ligand-receptor bond.
-barrel of the streptavidin protomer. Only the carboxylate
oxygens and the ureido-ring nitrogen protrude to the outside.
Multitudinous hydrogen-bonds and van der Waals interactions are involved
in the biotin binding. There are three hydrogen bonds to the carbonyl
group buried within the barrel and also hydrogen bonds to the ureido
nitrogens and carboxyl oxygens. Four tryptophan amino acids are in
contact with each biotin molecule. Most of the interactions result from
the residues of a given subunit, although one tryptophan is supplied by
a subunit related to the R diad axis, which is vital for the tetramer
integrity. This is an explanation for the reduced affinity with less
than four ligands that was also found elsewhere
[76,113]. WEBER et al. [131] confirmed
the results for streptavidin and apostreptavidin in 1992 and
LIVNAH et al. [88] got similar results for the
avidin-biotin complex in 1993. In 1998, CHU et al. [24]
presented a very convincing experiment, where they removed the
polypeptide loop that undergoes an open to closed conformational change
when biotin is bound. They showed that the deletion caused a large
decrease of the affinity for the full ligand-receptor bond.
Although JONES and KURZBAN [72] presented good evidence that the streptavidin-biotin binding is not cooperative1.5 in the sense of the MWC-model1.6, many other publications clearly indicate that the bond strength makes a step for the full bond. This can be seen as a positive cooperativity, as proposed by WILLIAMS et al. [133] and SANO et al. [113]. Besides the already mentioned x-ray diffraction experiments, these results were also supported by fourier-transform infrared spectroscopy and fluorescence spectroscopy [49].
0.5
![\includegraphics[width=.5\textwidth]{Bilder/two-well-potential}](img34.png)
|
All theoretical descriptions of ligand-receptor bonds and their
breaking is based on the Transition-State-
Theory (TST) and
KRAMERS' model [15]. The TST was developed
by
POLANYI and WIGNER in 1928
[103] and expanded by EYRING in 1935
[41]. Generally the classical TST describes any two
physical states that are separated by a bottleneck in
phase
space. Two assumptions have to be made to apply the TST. First, the
strong-coupling assumption, i.e. that all effects from a divergence of
the thermal equilibrium are neglected, and second, the point of no
return, i.e. that separated states do not reconnect. Even when these
assumptions are applicable, TST can only give an upper bound to the true
rate for any dividing surface [61]. KRAMERS'
model describes a chemical reaction as a classical particle that moves
in a one dimensional asymmetric double-well potential (confer to
figure 1.5). A thorough description of KRAMERS'
model can be found in [61].
The new experimental results about ligand-receptor bonds stimulated
several theoretical works that extend the TST and KRAMERS'
model. In 1996 GRUBMÜLLER et al. [59]
presented computer-simulations of the streptavidin-biotin bond that
matched their AFM measurements. The simulations supported their
experimental results, that the measured bond-force increased with an
increasing loading rate, and that the bond-force is around 280pN for a
loading-rate of over 100nN/sec. But they simulated an extremely stiff
cantilever (the spring constant was nearly 20 times higher than in the
experiments), the time-scale of force increase was nanoseconds rather
than milliseconds (as in experiments) and only a streptavidin monomer
has been simulated. In 1997 IZRAILEV et al. simulated the
avidin-biotin bond and found very high rupture forces of up to 450pN.
They also presented a theoretical study that demonstrates that the
nanosecond simulations can not reproduce thermally activated bond
rupture that requires milliseconds. Also in 1997 EVANS and
RITCHIE [40] published a thorough extension of
KRAMERS' model to simulate force-activated bond rupture, and
tested their model using smart Monte Carlo simulations. Additionally,
they proposed a law for the exponential loading-rate dependency of the
avidin-biotin rupture between 1 and 10![]() pN/sec and 100 to
400pN.
pN/sec and 100 to
400pN.
0.5
![\includegraphics[width=.5\textwidth]{Bilder/energy-landscape}](img36.png)
|
As shown 2000 by STRUNZ et al.
[121] the
simplest possible model for ligand-receptor pairs has at least one
intermediate state. Figure 1.6 illustrates such
a model. Without any outer force the width of the potential (![]() ) is
wide. An applied force (represented by the dashed line) lowers the outer
barrier and therefore the inner barrier with a narrower potential width
(
) is
wide. An applied force (represented by the dashed line) lowers the outer
barrier and therefore the inner barrier with a narrower potential width
(![]() ) is relevant for the bond rupture properties. For the narrowing
of the potential width a linear lever rule is valid: The smaller the
potential width, the higher the rupture force. These models can only be
applied if there is a single preferred path for the reaction
[15],
) is relevant for the bond rupture properties. For the narrowing
of the potential width a linear lever rule is valid: The smaller the
potential width, the higher the rupture force. These models can only be
applied if there is a single preferred path for the reaction
[15],
which was shown experimentally by
FREITAG et al. [45]. Such a model was introduced in
1978 by BELL [8] and used e.g. by MERKEL
et al. to explain the loading-rate dependency of the ligand-receptor bond
in 1999. Experimental data from bond-force measurements can be fitted
to a linear function [121]: